一、太古宙固氮微生物的矿物利用策略
仰望天空时,或许很少有人会想到,占据大气体积78%的氮气竟是地球生命所面临的一场“悖论盛宴”。氮元素是蛋白质和DNA的基石,在生命体系中不可或缺,然而其在大气中的主要存在形式——氮气(N2)却由于氮氮三键的巨大键能而表现出极强的化学惰性,对绝大多数生命形式关闭了直接利用的大门。将大气中的氮气转化为生命可同化的氨(NH3),需要突破化学界最强的化学键之一。完成这一关键步骤的,并非雷霆万钧的闪电或人类工业革命创造的哈伯-博施工艺,而是一群肉眼不可见的微小生命——固氮微生物。它们与地球环境之间的协同演化,构成了跨越数十亿年的宏大演化史诗。
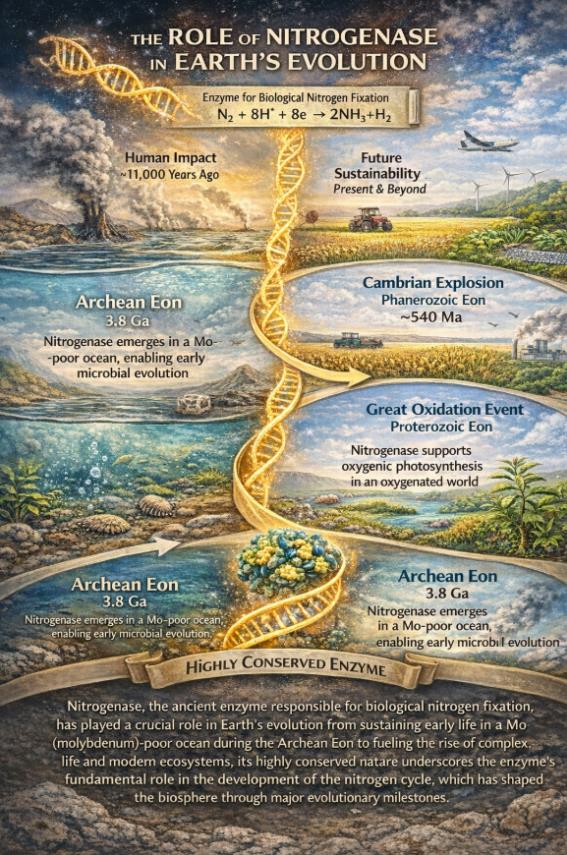
回溯至距今约38亿至25亿年前的太古宙,早期地球的海洋处于还原性状态,与现代海洋不同,其中溶解态的钼(Mo)极为匮乏。然而,钼恰恰是绝大多数高效固氮酶(钼铁蛋白)活性中心的核心金属辅因子。这就构成了一个关键的演化谜题:在缺钼的海洋环境中,早期生命究竟如何开启氮气利用的通道?中国地质大学(北京)的一项研究为这一问题的解答提供了重要线索。研究发现,一种名为沼泽红假单胞菌(Rhodopseudomonas palustris)的不产氧光合细菌,在厌氧环境下具备利用辉钼矿(MoS2)这类矿物作为唯一钼源的能力[1]。该微生物的策略极为精妙:一方面,它通过分泌一种被称为“Rhodopetrobactin”的金属载体,这种小分子化合物能够有效螯合并剥离矿物表面的钼离子;另一方面,电子显微镜观察显示,微生物细胞会紧密贴附于辉钼矿表面,从而实现高效的元素摄取。这一发现的意义在于,它揭示了太古宙贫瘠海洋中的早期固氮微生物并非被动地依赖溶解态营养盐,而是能够通过主动的矿物-微生物相互作用,从海底热液喷口周围广泛存在的硫化物矿物中获取稀缺的金属辅因子。正是这种能力,支撑了早期固氮作用的繁衍,进而扩大了原始生物圈的生产力,为后续产氧光合作用的累积乃至大氧化事件的发生奠定了基础[1,2]。
二、固氮酶作为“分子时间胶囊”的稳定性
如果说微生物从矿物中获取金属元素是生命演化的起点之一,那么固氮酶本身在漫长的地质年代中又经历了怎样的演化轨迹?令人惊奇的是,这种核心酶表现出“分子时间胶囊”的特征,忠实地记录了地球环境的变迁。根据发表于《自然-通讯》的一项突破性研究,科学家借助合成生物学技术,成功“复活”了距今约7亿年至23亿年前的古代固氮酶基因,并将这段远古DNA植入现代细菌中,观察其功能表达[3]。研究结果显示,尽管这些复活的远古酶在催化效率上与现代版本存在差异,但在固氮过程中所产生的氮同位素分馏效应(ε15N)却表现出惊人的稳定性,与现代微生物几乎没有差别。这一现象意味着,在地球历史上,无论是大氧化事件(约24亿年前)引发的环境剧变,还是随后“雪球地球”时期的极端严寒气候,固氮酶作为生命核心引擎的工作方式,即其同位素“指纹”,始终保持着高度保守性。这也解释了地质记录中的一个重要现象:在距今约32亿至27亿年的古老沉积岩中,氮同位素信号始终徘徊于0‰附近,指示着以钼固氮酶为主导的氮循环模式早已在地球上建立[4]。这些微小的工程师们,凭借其极其保守的化学机制,为地球生命支撑起了一张稳定而持久的营养安全网。
三、有氧环境下硝化与反硝化作用的形成
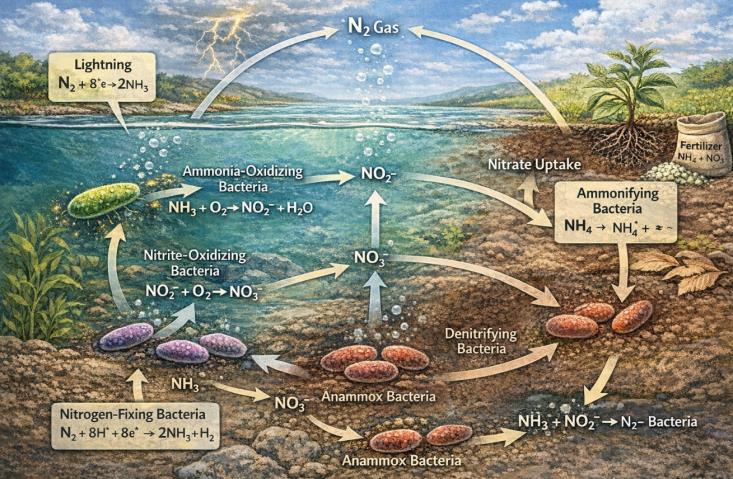
生命活动的持续演化反过来又深刻地重塑了地球环境。当产氧光合作用(主要由蓝细菌完成)大规模出现并积累,大气中的氧气含量急剧攀升,地球由此进入有氧时代。这一转变彻底重塑了全球氮循环的格局。在无氧条件下,氨(NH₃)是稳定的还原态氮素存在形式;然而,氧气的大量出现为一系列新型微生物代谢途径的演化创造了条件。化能自养微生物群落开始登上历史舞台,其中,氨氧化微生物(如奇古菌Thaumarchaeota)和亚硝酸盐氧化微生物(如Nitrospira)通过协同作用,将氨逐步转化为硝酸盐(NO3⁻),这一过程被称为硝化作用[2]。硝化作用产生的硝酸盐溶解性强、迁移性高,成为真核藻类及后续陆生植物最易于同化的氮源形式。与此同时,在缺氧的水体或沉积物环境中,另一类微生物则利用硝酸盐作为呼吸作用的电子受体,将其逐步还原为氮气(N2)并释放回大气,这一过程被称为反硝化作用[5]。至此,现代氮循环的完整闭环——包括固氮、硝化、反硝化作用——正式形成。这一闭环不仅是一系列生物化学反应的简单循环,更构成了地球系统的一种关键全球调控机制。如果没有反硝化作用将过量活性氮及时返回大气,海洋和陆地生态系统将因持续的氮富集而走向富营养化崩溃。
四、寒武纪生命大爆发时期的氮循环调控
在距今约5.4亿年前的寒武纪生命大爆发前夕,这种氮循环的调控机制表现得尤为突出。研究表明,在中国华南扬子地块的早寒武世地层中,硅酸盐结合态氮的同位素出现了异常偏高的记录,指示当时海洋环境中存在强烈的厌氧氨氧化(Anammox)过程[6]。厌氧氨氧化细菌能够在缺氧条件下将氨和亚硝酸盐直接转化为氮气,这是一种化能自养代谢途径。这一过程的存在意味着,在早寒武世广泛发育的缺氧海水中,微生物群落正在高效地处理氮素,从而调节着海洋中的活性氮储量。这种微生物代谢活动不仅直接参与调控氮循环的动态平衡,甚至可能通过影响碳同位素的波动(即SHICE事件),间接地与寒武纪生命大爆发这一重大演化事件产生关联[6,7]。
五、微生物代谢与地球环境的协同演化机制
回顾数十亿年的地球演化历程,不难得出一个基本结论:地球环境的变迁与微生物的代谢进化,始终是一部无法分割的协奏曲。在太古宙,面对缺钼海洋的营养限制,固氮微生物通过从岩石中“采矿”的策略,以顽强的化学手段开启了氮气资源的大门,支撑了早期生命系统的繁衍生息。进入元古宙,大氧化事件之后,好氧的硝化微生物迅速崛起,硝酸盐库的建立为真核藻类等复杂光合生物的繁荣提供了关键营养基础。到了显生宙,在大规模灭绝、大洋缺氧事件等剧烈气候扰动中,反硝化和厌氧氨氧化微生物则扮演了“安全阀”的调控角色,通过动态调节海洋活性氮储量,持续反馈并影响着气候与生物的演化轨迹。进入人类世,人类通过哈伯-博施工艺实现的工业固氮,其规模已远超所有自然陆地固氮的总和,从根本上改变了全球氮循环的自然格局。这一改变在支撑粮食产量大幅增长的同时,也引发了水体富营养化、温室气体N₂O排放加剧等严峻环境挑战。重新审视这段由微生物书写的地球演化史,不仅是为了满足对遥远过去的求知欲,更是为了在面对人类活动深刻改变地球系统的当下与未来,寻找到与这颗星球重新建立和谐共存关系的智慧与路径。直至今日,支撑整个地球生物圈可持续运转的,依然是那些微小工程师们数十亿年来一以贯之的古老代谢法则。
本文作者:厦门大学黄宇彬。本文由厦门大学海洋与地球学院焦念志院士、碳中和创新研究中心胡晨副研究员指导,由海洋负排放(ONCE)国际大科学计划、厦门大学碳中和创新研究中心支持。
参考文献
[1] Hou, L. F., Hao, J. L., et al. (2024). Molybdenite as a bioavailable molybdenum source for biological nitrogen fixation. Nature Communications, 15(1), 2345.
[2] Canfield, D. E., Glazer, A. N., & Falkowski, P. G. (2010). The evolution and future of Earth’s nitrogen cycle. Science, 330(6001), 192-196.
[3] Garcia, A. K., et al. (2020). Reconstructing the evolutionary history and ancestral states of nitrogenase. Nature Communications, 11(1), 1180.
[4] Stüeken, E. E., et al. (2015). Isotopic evidence for biological nitrogen fixation by molybdenum-nitrogenase from 3.2 Gyr. Nature, 520(7549), 666-669.
[5] Falkowski, P. G. (1997). Evolution of the nitrogen cycle and its influence on the biological sequestration of CO2in the ocean. Nature, 387(6630), 272-275.
[6] Wang, J. W., et al. (2020). Nitrogen isotope evidence for a perturbed nitrogen cycle during the SPICE event. Communications Earth & Environment.
[7] Zerkle, A. L., & Mikhail, S. (2017). The biological nitrogen cycle through geological time. Geobiology, 15(3), 343-360.
责编:微科普